
Saturday, 15 July 2006
138-76
A Model for Soil Organic Matter Turnover Proposed from Laboratory and Field Labelling Experiments and Sensitivity Analysis.
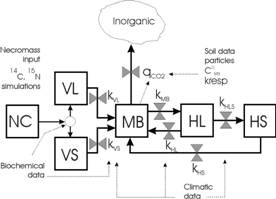
The natural or artificial isotopic tracers constitute an essential tool to understand and model the transformations of soil organic matter (SOM). Added into one of the organic compartments, then followed through the other compartments assumed to be homogeneous, the tracers provide an image of the functioning of the system. 14C and 15N labelled wheat straw was incubated in situ in fallow soils of Venezuelan Paramo. During two years the following data were collected: soil moisture, total-14C and -15N, microbial biomass MB-14C and -15N, rainfall, temperature and radiation. Soil moisture was daily predicted with the SAHEL model (Penning de Vries et al, 1989). For all the tested model alternatives, the added plant material (APM) was split into labile and stable plant compartments (Fig. 1) from APM biochemical composition according to equations proposed by Thuričs et al. (2001, 2002) from a range of contrasted organic materials. The aim of this work was a comparative exploration of flow diagrams of the other SOM compartments of microbial origin. Five alternatives to the MOMOS-1 model (Sallih and Pansu, 1993 ; Pansu et al., 1998) were tested to predict the flows of 14C and 15N tracers through these microbial compartments. These models were classified in two types. Classical models of type 1 (MOMOS-2 to -4) use two types of parameters: kinetic constants regulating the compartment outputs and partition factors regulating the compartment inputs (efficiency factors). The proposed models of type 2 (MOMOS-5 and -6) use only kinetic constants for compartment outputs. In type 2 models MB is the central compartment of the system: following the Jenkinson (1977) interrogation, MB can be considered as "the eye of the needle" through which all organic material must pass (Fig. 1).
MOMOS-2 was similar to MOMOS-1 for the C-cycle and included a simplification for the N cycle. MOMOS-3 was structurally similar to the Roth-C model of Jenkinson (1990). It was a simplification of MOMOS-2 which eliminated a compartment of labile metabolite (not measured in the field experiment). MOMOS-4 was a further simplification of MOMOS-3 which eliminated recycling of transformed materials from MB and humus compartments. MOMOS-5 explored two new modifications: (1) the whole of outputs from plant material and humus compartments was the MB input; (2) MB output was defined by the metabolic quotient for CO2 (qCO2, Anderson and Domsch, 1993) and MB mortality rate (kMB). MOMOS-6 complemented MOMOS-5 by introducing a stable humus compartment for long term predictions (HS in Fig. 1).
The analysis of accuracy and sensitivity (Pansu et al., 2004) showed: (1) the simplifications from MOMOS-2 to MOMOS-3 then from MOMOS-3 to MOMOS-4 were valid, (2) predictions of all tested models were not significantly different for total-14C and -15N, (3) the type 2 models significantly improved predictions of MB-14C and -15N showing a dynamics different of the predictions of type 1 model, (4) predictions by type 2 models were more robust than predictions by type 1 models in spite of a more complex formulation for MB description. Type 2 models presented also a higher ecological consistency (1) between predicted variations of MB activities and mineralization rates, (2) between links of parameter fluctuations and model responses. The improvement brought by MOMOS-6 was not significant compared to MOMOS-5 during the two year experiment, but allowed to quantify 14C and 15N sequestered in HS. This comparative study enables to recommend the MOMOS-6 concept as a basis to simulate the dynamics of both native OM and fresh materials which are yearly brought to the soil.
Fig. 1 – The selected MOMOS-6 model from (1) results of laboratory experiments (NC, VL and VS), (2) results of field labelling experiment and sensitivity analysis (MB, HL and HS).
Back to 2.2A Soil Organic Matter: Stabilization and Carbon Sequestration - Poster
Back to WCSS
Back to The 18th World Congress of Soil Science (July 9-15, 2006)